進行中の研究
試験管内筋層培養システムも構築と大腸癌浸潤機構の解明
大腸癌は粘膜で発生し、増殖し、筋層にて浸潤します。これまでマトリゲルなどを用いて浸潤機構が研究されてきましたが、生体内の浸潤の場である筋層で、癌細胞はどのように振舞うのか、観察した報告はありません。生理学の分野では筋層培養は汎用されています。この技術を活用し、癌細胞の浸潤機構の解明を目指します。
大腸癌の肝転移を決める因子の同定
大腸癌は血行性に肝臓に転移することが知られています。マウス由来の大腸癌細胞を同系のマウスに移植することで肝臓によく転移する細胞群を得ることが出来ました。肝臓へ転移する細胞特異的なその分子機構は何か?探っていきます。
乳癌の肝転移を決める因子の同定
乳癌でも数は少ないですが、肝転移を起こします。その機構は大腸癌と同じなのか、乳癌特異的なのか。調べていきます。
終了した研究
大腸癌の肝転移にはコレステロールが必要である
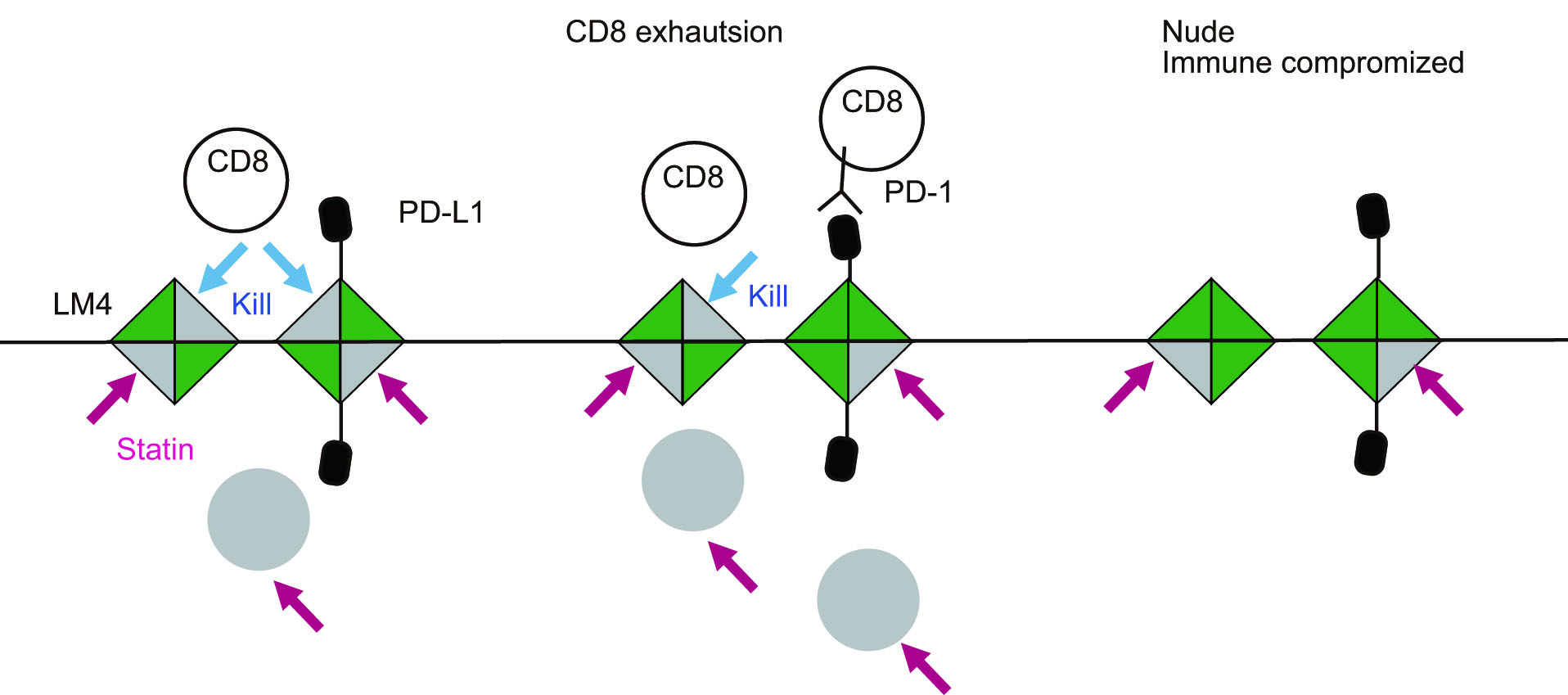 大腸癌患者の半数は肝転移を示します。肝転移特異的な遺伝子変異は知られておらず、どのような細胞が肝転移に適しているか、まだわかっていません。Colon26(KRas活性化変異がある)をBALB/cマウスの脾臓に打ちこんで、肝臓に転移した細胞を取ってきて、また脾臓に打つ、というサイクルを4回繰り返し、LM4細胞を取得しました。C26とLM4と比較すると、血管外遊走能(三浦先生の作った試験管内血管で観察)には差がなかったのですが、代謝が異なっていることがわかりました。グルコースをよく使うC26に対し、LM4は脂肪酸を取り込み、コレステロールを良く作っていることがわかりました。コレステロール合成阻害薬であるスタチン処理でLM4は増殖が抑制されました。肝転移したマウスにスタチンを処理すると、野生型マウスでは効果が得られなかったのに対し、ヌードマウスでは効果が認められました。これは、野生型マウスよりもヌードマウスの方がLM4が育ちやすいことに起因するものです。スタチンはマウスのCD8T細胞を選択的に死滅させる報告が以前にあることから、図のようなモデルを考えています。コレステロールがあるとどうして肝でよく育つのかは今後の課題です。九大・三浦研、金大・高橋研との共同研究です。Taniguchi, Hoshi & Kiyokawa, Can Sci., 2024.
大腸癌患者の半数は肝転移を示します。肝転移特異的な遺伝子変異は知られておらず、どのような細胞が肝転移に適しているか、まだわかっていません。Colon26(KRas活性化変異がある)をBALB/cマウスの脾臓に打ちこんで、肝臓に転移した細胞を取ってきて、また脾臓に打つ、というサイクルを4回繰り返し、LM4細胞を取得しました。C26とLM4と比較すると、血管外遊走能(三浦先生の作った試験管内血管で観察)には差がなかったのですが、代謝が異なっていることがわかりました。グルコースをよく使うC26に対し、LM4は脂肪酸を取り込み、コレステロールを良く作っていることがわかりました。コレステロール合成阻害薬であるスタチン処理でLM4は増殖が抑制されました。肝転移したマウスにスタチンを処理すると、野生型マウスでは効果が得られなかったのに対し、ヌードマウスでは効果が認められました。これは、野生型マウスよりもヌードマウスの方がLM4が育ちやすいことに起因するものです。スタチンはマウスのCD8T細胞を選択的に死滅させる報告が以前にあることから、図のようなモデルを考えています。コレステロールがあるとどうして肝でよく育つのかは今後の課題です。九大・三浦研、金大・高橋研との共同研究です。Taniguchi, Hoshi & Kiyokawa, Can Sci., 2024.
連続切片免疫染色の定量化による、浸潤端にある大腸癌細胞の蛋白質発現と予後
 浸潤端にある大腸癌細胞は浸潤・転移を担うと考えられています。この細胞群はどのような性質であるのか、免疫染色で検討されてきましたが、同一細胞での発現解析報告は少数です。連続切片を用いても同じ部分を探すのは時間がかかるという問題があります。
そこで、QuPathによって複数の免疫染色像を同時に動かし、既存のライブラリーを活用し、Pythonにて画像を回転させ、半自動で同じ部位を見つける方法を確立しました。
免疫染色画像を2値化し、浸潤端での癌細胞での発現を調べると、p16の発現が高くKi67の発現が低い、あるいは、p21の発現が低く、Ki67の発現が低い群は予後が悪いことがわかりました。Go or Grow仮説は、細胞は移動するか・増殖するかのどちらかを選択しているという説ですが、p16やp21は老化マーカー、Ki67は増殖マーカーなので、我々の結果はそれを支持しているといえます。但し、同じ老化マーカーでもp16は高く、p21は低いのはどう説明するのか?
p16は細胞質にも発現し、細胞骨格を制御するという報告があります。細胞周期を止める以外の機能がプラスされた細胞ががんの浸潤転移を担うのかもしれません。一般消化器外科との共同研究です。Kin, Hoshi & Kiyokawa, Pathol. Int, 2023.
浸潤端にある大腸癌細胞は浸潤・転移を担うと考えられています。この細胞群はどのような性質であるのか、免疫染色で検討されてきましたが、同一細胞での発現解析報告は少数です。連続切片を用いても同じ部分を探すのは時間がかかるという問題があります。
そこで、QuPathによって複数の免疫染色像を同時に動かし、既存のライブラリーを活用し、Pythonにて画像を回転させ、半自動で同じ部位を見つける方法を確立しました。
免疫染色画像を2値化し、浸潤端での癌細胞での発現を調べると、p16の発現が高くKi67の発現が低い、あるいは、p21の発現が低く、Ki67の発現が低い群は予後が悪いことがわかりました。Go or Grow仮説は、細胞は移動するか・増殖するかのどちらかを選択しているという説ですが、p16やp21は老化マーカー、Ki67は増殖マーカーなので、我々の結果はそれを支持しているといえます。但し、同じ老化マーカーでもp16は高く、p21は低いのはどう説明するのか?
p16は細胞質にも発現し、細胞骨格を制御するという報告があります。細胞周期を止める以外の機能がプラスされた細胞ががんの浸潤転移を担うのかもしれません。一般消化器外科との共同研究です。Kin, Hoshi & Kiyokawa, Pathol. Int, 2023.
細胞融合を制御する機構

生体内で細胞は融合します。代表的なものとして、筋芽細胞、マクロファージ、癌細胞、胎盤の栄養膜細胞が知られています。DOCKファミリーはショウジョウバエの筋芽細胞の融合に必須な分子として単離されましたが、他の細胞種の融合に関与するかは明らかではありませんでした。胎盤由来の栄養膜細胞での発現をみると、DOCK1とDOCK5が発現していました。それぞれに特異的な阻害薬を投与すると、細胞の融合は阻害されるのではなく、促進されました。直接的にDOCKファミリーが融合過程に効くというよりは、様々な遺伝子発現を変化させることによって融合を阻害しているようです。 また、DOCK1とDOCK5の機能は異なることも示唆されました。本学生物学の東海林先生、金大の大黒先生との共同研究です。TBOPPを供与頂いた九大・福井先生に感謝します!Kiyokawa, BBRC., 2020.
大腸癌の浸潤端にあるPDCの意義・発生機構の解明
 大腸癌ではBuddingあるいはPoorly Differentiated Cluster (PDC)という癌細胞からなる細胞塊があると予後が悪い(=遠隔臓器に転移する)ことが知られていますが、これらの細胞塊はどのように形成され、どのように生体で振る舞うのか?は明らかにされていません。
ヒト手術材料をEzrinで免疫染色すると、PDCの隅にある細胞でEzrinを発現している像が多いことに気が付きました。これを定量化する方法を確立しEzrin Corner Score(ECS)と名付け、ECSが高いと予後が悪いことを見出しました。HEでは同様に見えるPDCのなかでも蛋白質の発現が不均一で悪性度の高いPDCがあることを示唆するものです。本学・一般消化器外科との共同研究です。Aikawa & Kiyokawa, Can Sci., 2019.
大腸癌ではBuddingあるいはPoorly Differentiated Cluster (PDC)という癌細胞からなる細胞塊があると予後が悪い(=遠隔臓器に転移する)ことが知られていますが、これらの細胞塊はどのように形成され、どのように生体で振る舞うのか?は明らかにされていません。
ヒト手術材料をEzrinで免疫染色すると、PDCの隅にある細胞でEzrinを発現している像が多いことに気が付きました。これを定量化する方法を確立しEzrin Corner Score(ECS)と名付け、ECSが高いと予後が悪いことを見出しました。HEでは同様に見えるPDCのなかでも蛋白質の発現が不均一で悪性度の高いPDCがあることを示唆するものです。本学・一般消化器外科との共同研究です。Aikawa & Kiyokawa, Can Sci., 2019.
腺管の回転と癌の浸潤・転移
 MDCK細胞はイヌ尿細管由来の細胞で、通常の培養皿にまけば自発的に遊走しますが、細胞外マトリックスに富んだ環境では細胞塊や類器官と呼ばれる3次元構造を形成します。この構造は生体内の腺管(あるいは腺房)構造を模していると考えられています。この構造に膵癌や大腸癌で変異の見られるK-Rasの恒常的活性化型を発現させると回転することを見出しました。
細胞塊では腺管よりも回転速度が大きいことがわかりました。また細胞塊ではCCNB1が必要ですが、腺管ではHDAC阻害薬処理や活性型b-カテニンの発現によって回転が促進することを見出しました。腺管の回転を促進する因子は細胞塊の回転では無効であることも明らかにし、分子機構が異なることも示唆されました。試験管内で観察された回転が生体内でも見られることも考えられますし、間質に様々な因子や細胞がある状態では異なるフェノタイプとして表現されているかもしれません。
今後は生体内で細胞塊や腺管はどのような振る舞いをするのか、明らかにしていきたいと思います。金沢大学の堀家先生にアレイを行って頂きました。Hirata, Ichikawa, & Kiyokawa, Can Sci., 2018.
MDCK細胞はイヌ尿細管由来の細胞で、通常の培養皿にまけば自発的に遊走しますが、細胞外マトリックスに富んだ環境では細胞塊や類器官と呼ばれる3次元構造を形成します。この構造は生体内の腺管(あるいは腺房)構造を模していると考えられています。この構造に膵癌や大腸癌で変異の見られるK-Rasの恒常的活性化型を発現させると回転することを見出しました。
細胞塊では腺管よりも回転速度が大きいことがわかりました。また細胞塊ではCCNB1が必要ですが、腺管ではHDAC阻害薬処理や活性型b-カテニンの発現によって回転が促進することを見出しました。腺管の回転を促進する因子は細胞塊の回転では無効であることも明らかにし、分子機構が異なることも示唆されました。試験管内で観察された回転が生体内でも見られることも考えられますし、間質に様々な因子や細胞がある状態では異なるフェノタイプとして表現されているかもしれません。
今後は生体内で細胞塊や腺管はどのような振る舞いをするのか、明らかにしていきたいと思います。金沢大学の堀家先生にアレイを行って頂きました。Hirata, Ichikawa, & Kiyokawa, Can Sci., 2018.
レンズ上皮細胞におけるDOCK5発現欠損の影響
 DOCK5蛋白質は欠損により、レンズ核が後極に逸脱することが知られていますが、その機序は不明でした。逸脱前の3週齢のマウスのRNAからマイクロアレイを抽出しIPA解析をすることでDOCK5欠損によって引き起こされる細胞内伝達経路を推定しました。本学眼科との共同研究です。Xu, Yoshizaki, & Kiyokawa, Mol Vis., 2017.
DOCK5蛋白質は欠損により、レンズ核が後極に逸脱することが知られていますが、その機序は不明でした。逸脱前の3週齢のマウスのRNAからマイクロアレイを抽出しIPA解析をすることでDOCK5欠損によって引き起こされる細胞内伝達経路を推定しました。本学眼科との共同研究です。Xu, Yoshizaki, & Kiyokawa, Mol Vis., 2017.
上皮再生の生体内イメージング
 虚血性腸炎腸では虚血によって壊死が起こることが知られています。カプセル型内視鏡の発達・普及により、大腸だけでなく小腸でも虚血性腸炎が起こっていることがわかってきました。マウスと2光子顕微鏡を用いて、上皮の壊死・脱落から上皮が再生される分子の仕組みをイメージングにより解明しました。Takeda & Kiyokawa, Sci Rep., 2017
虚血性腸炎腸では虚血によって壊死が起こることが知られています。カプセル型内視鏡の発達・普及により、大腸だけでなく小腸でも虚血性腸炎が起こっていることがわかってきました。マウスと2光子顕微鏡を用いて、上皮の壊死・脱落から上皮が再生される分子の仕組みをイメージングにより解明しました。Takeda & Kiyokawa, Sci Rep., 2017
MDCKオルガノイドの脂質解析
 MDCK細胞のオルガノイド(シストあるいは類器官)は上皮の生体内3次元構造を模する系として活用されていますが、その際の脂質構成については不明でした。2次元vs3次元、3次元の早期と後期、正常と癌(K-Ras変異体の導入)の各々の状態について、質量分析とマイクロアレイを用いて解析しました。本学、血液内科・岡崎俊朗グループとの共同研究です。Yoshizaki & Kiyokawa, J Biochem, 2016
MDCK細胞のオルガノイド(シストあるいは類器官)は上皮の生体内3次元構造を模する系として活用されていますが、その際の脂質構成については不明でした。2次元vs3次元、3次元の早期と後期、正常と癌(K-Ras変異体の導入)の各々の状態について、質量分析とマイクロアレイを用いて解析しました。本学、血液内科・岡崎俊朗グループとの共同研究です。Yoshizaki & Kiyokawa, J Biochem, 2016
MDCKオルガノイドの成長過程で発現が上昇するRipply1の分解機構
 MDCK細胞のオルガノイドが形成される際の早期と後期に着目して、後期で発現が上昇してくるRipply1の細胞内動態について解析しました。市販の抗体では内因性の蛋白質を染色することが出来ず、培養細胞のみの解析で終わってしまったのが残念です。学部生2年生だった桑島さんが実験の立ち上げをしてくれました。Yoshizaki, Kuwajima & Kiyokawa, BBRC., 2015
MDCK細胞のオルガノイドが形成される際の早期と後期に着目して、後期で発現が上昇してくるRipply1の細胞内動態について解析しました。市販の抗体では内因性の蛋白質を染色することが出来ず、培養細胞のみの解析で終わってしまったのが残念です。学部生2年生だった桑島さんが実験の立ち上げをしてくれました。Yoshizaki, Kuwajima & Kiyokawa, BBRC., 2015